
Family Tree DNA held their 11th International Conference of Genetic Genealogy from November 13-15, 2015 in Houston, Texas.

First and foremost, I want to thank Max Blankfeld and Bennett Greenspan, founders of Family Tree DNA, for hosting and subsidizing this conference. It’s the only conference of its type anyplace in the world and Family Tree DNA has been hosting this conference now for 11 years.
Now to the fun part – the conference itself.
The first year I attended, which was the first conference in 2004, I remember thinking and probably saying as well that I felt like I was drinking from a firehose. It reminded me of grad school.
This was both the good news and the bad news. The good news was that I loved every minute. The bad news is that I didn’t understand everything that was being said. And there was one session in particular where I was sure I had wandered into the wrong conference room…but when I saw Max and Bennett, I was sure I was in the right room….but not at all sure I was in the right place.
However, we were all newbies together because the field of genetic genealogy didn’t exist until about the year 2000….so these were truly first baby steps – although they certainly didn’t seem like it at the time.
Fast forward to this year. Thanks to technology and the leaders in this field, the edge of the envelope is still being pushed, and there are still very exciting topics on the agenda for those of us who know a bit more now. Those topics didn’t even exist in 2004.
Looking at the 2015 agenda, there were 7 main sessions, plus the breakouts and lab updates and such. Of those 7 main sessions, none of them, not one, could have been presented at the conference just 5 years ago. Why? Because the products and the tools didn’t exist then. We have advanced a HUGE distance in just 5 years and much of this has been due to citizen scientists.
But not every conference session was at an advanced level. Thanks to Max and Bennett, there were also breakout sessions for newbies who I’m sure, feel like they are drowning in that fire hose.
There were 206 attendees at this year’s conference, and of those, I think there were 70 first-timers. I surely hope they come back, because it does get easier as you learn more about the topics at hand.
The conference always begins with a reception hosted by Family Tree DNA on Friday evening, the ISOGG party (where we all contribute food and beverages) on Saturday evening and whoever is left on Sunday evening tends to gather and eat the nonperishable leftovers from Saturday’s gathering.
Indeed, we have fun and visit from the time we get up in the morning until we close our eyes each evening. We take advantage of every minute.
Dressing the Part and Sharing the Love
You can tell you’re in a group of genetic genealogists. Just look at their clothes – Katherine Borges and Linda Magellan’s clothes in particular. They could start a helix clothing line!

But it doesn’t end there.

Look at the carpet too.
Not only that, I’m sure Katherine’s nail tech is sticking pins in a voodoo doll of Katherine.

Katherine’s break-out session was titled, “Nulls, the Value of Nothing” and she had named her nails 389, 425, 464, etc. for the markers that sometimes have null values.
I want to infect you with some of the rejuvenation and excitement we feel, especially those of us returning year after year. It’s how we charge our genetic batteries.
The conference, aside from providing us with an incredible learning opportunity, provides us with the opportunity to network and to visit with each other. There is indeed a lot of shop-talk going on…but there is also a lot of just plain visiting and laughing and fun. It’s kind of like a cross between a class reunion, a professional academic conference and a family reunion. And really, it’s the only place you can go and have these kinds of discussions. I mean truly, your family does not want to hear about this over the Thanksgiving table. But your genetic genealogy friends do!
Jennifer Zinck did us all a huge favor and took exceptional notes during the conference in the sessions she attended. Her posts are at these two links.
http://www.ancestorcentral.com/11th-international-conference-on-genetic-genealogy/
http://www.ancestorcentral.com/11th-international-conference-on-genetic-genealogy-sunday/
Due to the internet speed in the hotel, Jennifer was not able to upload any photos. I’m not about to recreate all of Jenn’s hard work, so what I am going to do is share some photos and what I considered to be salient and high points of the conference.
Now for the bad news, my camera battery ran out at the end. I thought I had an extra one with me, but I didn’t, so I don’t have photos of every main session, nor of the breakouts.
Update: Please note that the speaker’s slides are available here.
Welcome

Max and Bennett always open the conference with some comments and a welcome. Bennett shared a story I never knew about him. When Bennett was 13 years old, he drew his first genealogy pedigree chart after talking to his grandparents and older family members. While that is remarkable enough, he was bright enough to draw it – IN PENCIL. I’m still not bright enough to do that apparently.

If you ever wonder if the cosmos has a sense of humor, consider that Bennett has paid to test 68 Greenspan men and none of those 68 men have been a match to his line. However, 15 years into this adventure, a Mr. Green approaches Bennett at a conference and wonders if they are related. Bennett is tired of paying for unproductive tests and really doesn’t think there is ANY chance of Mr. Green matching him. So, Mr. Green pays for his test and you know what’s coming don’t you….yep….Mr. Green is Bennett’s closest match….and Mr. Green has a village name in the old country. I’m betting Bennett is going to be going on a trip soon to that village….don’t you! Maybe Mr. Green will go along. I’m extremely glad Bennett is finally reaping the harvest of his infinite patience.
Peter Sjolund – Y DNA Maps Scandinavian Family Trees from Medieval Times and the Viking Age

The first presentation was Peter Sjolund from Sweden.

The Scandinavians have become extremely excited about genetic genealogy and have been very active in projects and testing, including Big Y tests.
The first thing that happened was that I became exceedingly envious that the Hersesson family Peter was discussing in his presentation has 18 generations of documented family.
The great question to be answered was whether or not the farmer family was genetically related to the noble family.

The descendants were able to find enough direct male descendants to Y DNA test, and the answer was no, they weren’t. However, they discovered that STR markers just didn’t reach far enough back in time to provide matching and delineation or the farmer line who did match each other, so they added the Big Y test and managed to prove the oral history from 1350 to present.
In essence, Peter was building the family tree with SNP and STR data instead of records and the two sources confirm each other.
You know those stories about “there were two brothers – one went east and one went west?” Well, it’s true in this case, and using SNPs from the Big Y, Peter was able to prove it.
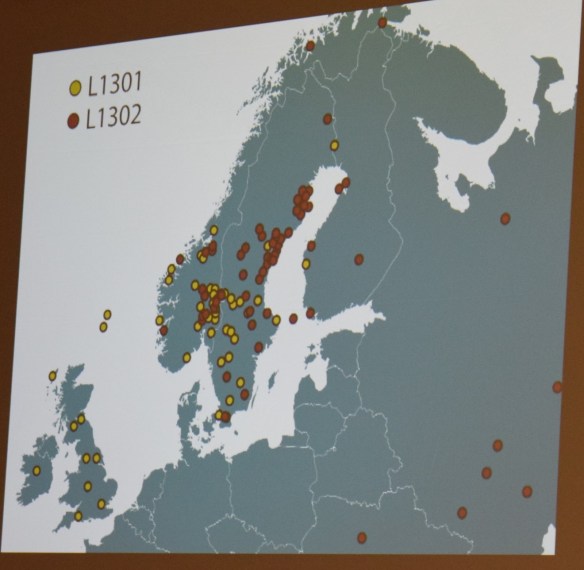
Peter’s next slide shows the historical events that spread these SNPs.
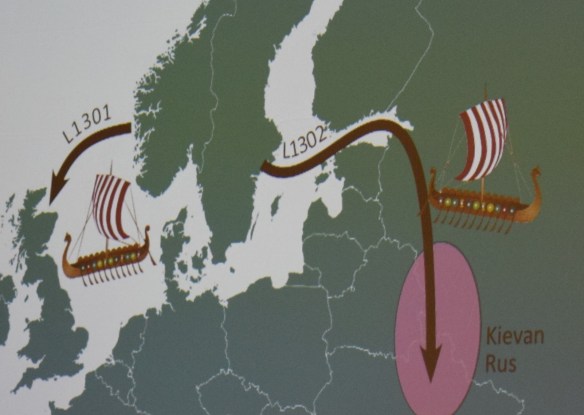
It was so enlightening to see exactly how this worked to prove the families, but also to connect to ancient history. Max summed it up well at the end when he said that “testing is not only a contribution to family history but also to confirming history itself.”
Razib Khan – Populations in Autosomal DNA
Razib, a doctoral candidate studying evolutionary genomics at UC Davis has been working with Family Tree DNA on updating their myOrigins product.

I loved Razib’s comment that all of this would have been considered science fiction 15 years ago. He’s absolutely right.
Razib described our autosomal ethnicity as being a rich diaspora and that it has to be measured, matched and then reduced to 50 narrative threads. He said it also gets a bit messy sometimes, because if we don’t fit into a thread exactly, or maybe our correct thread doesn’t have a deep enough reference population, we’re forced into the next best genetic thread – even though it may make no sense to us today. That does explain some of the odd results we see from time to time.

In order to improve myOrigins, the next version, due out in mid-late first quarter of 2016 will include several new reference populations, including a second reference population for Native American people.

Not all of Razib’s presentation was about ethnicity – some was about recombination – which of course, when you think about it, affects ethnicity dramatically.
He mentioned, almost in passing that in each meiosis, a male has 25 recombinations and a female, 35. This has the potential to affect the amount of autosomal DNA that we inherit through an all-female line of ancestors, for example, as compared to an all-male lineage. This means, in essence, that we are likely, over time, to carry more of our male lineages, or lineages heavily male, than we are do all female lineages – because the DNA divides less. I have to wonder if this is built into any of the calculations for relatedness by any of the vendors?
Razib mentioned that based on the results of ancient genome sequencing that the people of ancient Europe looked much different than the Europeans of today.
Razib also mentioned at least three instances where a combination of ancient DNA sequencing, population genetics and oral history have, together, proven the oral history to be accurate. One of these instances is the Aboriginal oral stories of the tribes in Australia that recall an Australia with a very different shoreline than today’s continent.

In the slide above, the light tan areas are now underwater, but the Aboriginal people still carry stories about these areas that have been “discovered” underwater.

Another “myth” is of giants in Biblical times. Recently one of the remains excavated was nearly 7 feet tall – a person who would surely have been considered a giant among men of that time.
Razib also talked a bit about full genome sequencing and a few other speakers touched on it as well. In essence there are four issues relative to full genome sequencing for the consumer marketplace.
- The cost of sequencing itself.
- The current lack of and cost of developing tools to compare full genomes.
- The knowledge of how to utilize the comparative results in a genetic genealogy context.
- The lack of any comparative database of other people.
Yes, I know that the other forms of DNA testing also started out with no data base, but those tests didn’t cost thousands of dollars either. So, in a nutshell, the technology to reduce the cost of the test itself hasn’t reached the level at which the consumer marketspace would embrace that testing. That’s probably when the work will be done on the tools, if at all. We really don’t know that more, in this case, full genome sequencing, would be enough better to warrant additional testing and development.
Razib closed by wondering if we will be able one day to “recreate the face of our ancestor” by utilizing the combined DNA of their descendants.

I have to admit, this would be VERY cool. It will be interesting to see what the next decade brings us in terms of technological advances.
I know one thing, if one had to do the “rebuild” by hand, the way I had to do the spreadsheet for James Crumley born in 1712, there won’t be very many faces recreated. Hopefully, by then, we’ll have better tools.
Razib’s memorable comment: “Treasure your exceptions.”
Dr. Michael Hammer – R1b and the Peopling of Europe: an Ancient DNA Update

I could hardly wait for this presentation by Michael Hammer.

For the past two years, Michael has updated us on the cumulative finds of ancient DNA, and translated or speculated upon what that means or will mean. Michael says that ancient DNA has changed the way we think of human origins and it will continue to do so in the future. I think that’s one the most dramatic understatements I’ve ever heard.
He also mentioned that humans incur about 70 mutations throughout their genome in each generation.
Michael went on to remind us that just because we find a population, as defined by a SNP, in very high numbers in a location today, doesn’t mean that is the origin of the population. The best example is that because R1b is found in about half of the European males today, it was long assumed that R1b was birthed in Europe – but it wasn’t.

Then Michael dropped the bomb on us – R1b is ANE and specifically is found among the Yamnaya. We had discussed this possibility last year, because no R1b is found in the earliest hunter-gatherer ancient remains in Europe. Subsequent research proved it. R1b comes from the Russian Steppes as is proven in the Haaks paper published in June 2015.
Today, 10 ancient Yamnaya samples have been analyzed, and all 10 are R1b. Hmmmm….
I wrote about the Yamnaya here.
One of the factors that has helped immensely with this problem is that in 2014, there had been about 30 ancient DNA samples sequenced and in 2014, there had been less than 80 sequenced. Today that number is at 160 and unexpected revelations are occurring.
We’ve known there were two populations for a long time that settled in Europe, the original-hunter gatherers and the farmers, but we didn’t know about a third population until relatively recently. Ironically, the day after the conference, word of a 4th population, from the Caucasus, broke.

The last group anyone expected R1b to emerge from was the ANE.

This map shows the influx of various cultures into Europe, and when.

Which haplogroups arrived when?

Oh, you wanted SNPs? Ok, here goes!

One word of caution from Michael is that when reading papers, understand that they may not always be comparing apples and apples. For example, the reason one SNP may not be present in a paper is not because it’s absent in the population or that particular sample, but because that lab for whatever reason, didn’t test for it. So, no assuming nor drawing non-match inferences allowed.
I had discovered this recently when reading a mitochondrial paper. They only tested for select locations and not others. Makes me absolutely crazy.
BreakOut Sessions
Emily Aulicino talked about “Supercharging Your Project Members.”
Jim Brewster talked about “Getting Started with GAP.”
Dr. Doron Behar discussed “The Origin of Ashkenazi Levites.”
I attended Dr. Behar’s lecture.

Doron’s focus for many years has been on the Jewish population.

Recently, he has re-evaluated the available data and used new Big Y data in order to attempt to define the source of the Levite population.

I particularly like this slide, below, because it so succinctly illustrates the difference between traditional Sanger sequencing which is what is being done when you take the traditional 12, 25, 37, 67 and 111 markers tests, and the next generation sequencing like the Big Y.

Sanger sequencing is illustrated on the bottom. Reactions at specific locations are measured and then analyzed by humans and then recorded to reflect a specific value.
Next generation sequencing utilizes scanning. In the top part of the slide, you can see several scans of each area. The quadrant on the left, if I’m counting right, had 27 scans of the same area. This is called coverage. The scans then are recorded and they have to be aligned. As you can see, the start and stop locations are not the same. Then the results at a particular location are counted. In this case, the dark squares show the same value in a particular location. So not all of the scans show the same thing. Of course, the value is compared to a reference chart for what is “normal” and then the variant values are recorded. Assuming in this case that the variant values are colored dark blue, 18 recorded the same variant value but one is misaligned.
So, if you’re wondering why there is so much discussion about read coverage, alignment and valid results in Big Y tests…this is it in a nutshell. Not to mention, as shown in the upper right quadrant, sometimes that location doesn’t read at all, so we have what are called “no-calls” to muck up the gears. Family Tree DNA has to decide what is a “valid” result when they return results to customers. Not everyone agrees with that threshold, so some people and groups do their own analysis. What really is valid? We don’t know for sure but the reason that the ISOGG tree requires Sanger sequencing before adding a new SNP location to a tree is to verify that the Big Y scans are accurate.
Doron designed a study utilizing both STR and SNP markers found in the Big Y to discover additional information about the source of the Levite population.

Based on several samples both within and outside of the known Jewish community, plus one family, the Horowitz, whose genealogy reaches far back into Jewish history, Doron was able to confirm that the Levite population did arise in the Near East.

I loved Doron’s comment, “The role of a scientist is to doubt.”
I would add that’s also the role of a genealogist and in particular, a genetic genealogist.
But Doron said something else EXTREMELY profound and I was extremely heartened to hear it.
“I was wrong.”
Yep, that’s what he said. Let me tell you why I found this so inspiring and encouraging.
In the academic community, researchers are encouraged to research and publish their findings, along, of course, with their research data and the reasons why they reached their conclusion.
In the future, new information or technology may become available, and that original information may need to be adjusted, corrected, or it may be outright wrong.
There is NO SHAME in being wrong. The only people who are never wrong are the people who do nothing. Thomas Edison’s lab was filled with many “failures,” all of which were learning exercises that led to success.
In fact, the bigger shame is in not publishing and keeping your data and discoveries to yourself. If you wait until you know “exactly,” that will never happen and you’ll never publish anything. Even if the information turns out to be incorrect, it’s still a foundation for future research.
If another academic disagrees with a paper, they don’t publicly berate their colleague as “incompetent”, suggest they are suffering from “ascertainment bias,” state that the global “we” don’t approve of their research methods, nor do they say their research is a folly – at least not in a public forum. If they have something to say, they are expected to do so with professional decorum and write a rebutting paper to share their own research and information as to why they disagree and how the data proves their point.
As has been proven, sometimes papers, especially early papers, are simply incorrect.
Often, as in Dr. Behar’s case, the original researcher, due to their high level of interest in the topic at hand makes additional discoveries that refutes or adjusts their earlier work.
Again, open research is encouraged.
I am hopeful that the genetic genealogy community can act within the same professional decorum standards. Participating in character assassination of those engaging in research discourages open sharing, discourages research and assuredly discourages new people from participating. No one wants to become a target.
In a professional setting, people disagree with ideas but remain friends. Disagreements aren’t personal attacks. There are no DNA police.
I am very encouraged not only by Dr. Behar’s work, but by his professional demeanor as well. Doron said that we all hold important information relevant to these discoveries. He’s right, and the way to free that information is to both test and continue to question, research and publish.
Miguel Vilar – Genographic 2015: New Markers, New Geno Kit and Accessing the Data Base

Dr. Miguel Vilar is the Science Manager for National Geographic’s Genographic project as well as a molecular anthropologist. He attended the Family Tree DNA conference in 2013 and we were very pleased to have him back this year to update us on the Genographic Project, now celebrating its 10th anniversary. It has been an incredibly exciting decade.
Ten years ago, the Family Tree DNA Conference was at the National Geographic headquarters in Washington DC to celebrate the FTDNA/Nat Geo collaboration.

The Genographic Project has been very successful with over 670,000 public participants.

In the beginning, the Genographic test was either a basic mtDNA test for females, or a 12 markers Y test for males. The Geno 2.0 test changed dramatically, and the new Geno 2.0 Next Generation test offers even more.
However, the Geno 2.0 Next Generation test isn’t either the 2.0 test nor next generation sequencing. So don’t get confused by the name.

For someone who has already taken all of these tests, there is no incentive to test again, but for a new person who wants a base mtDNA haplogroup, a Y haplogroup, ethnicity, autosomal results and to transfer into Family Tree DNA for autosomal matching – the $149 price tag is certainly a good value and it’s a great starting point. The unlock price at FTDNA will be $39, same as for Ancestry or the 23andMe V3 chip.
After transferring to Family Tree DNA, males can test the Y STR markers and they will already have over 17,000 haplogroup defining SNPs tested. Bennett said the SNPs known about a year ago when the cutoff for the new chip was made were included.
The new Geno 2.0 NG chip is an Illumina chip, customized but compatible with the chip used by Family Tree DNA.
National Geographic wants to expand their research partnerships as well to include qualified genetic genealogists, citizen scientists and those in other academic fields.

Nat Geo has established an application process. If you are interested, contact Dr. Vilar at his e-mail above.
Additionally, National Geographic has 11 new grantees doing fieldwork now in Chilean Patagonia.

An exciting aspect of this work is that 48 ancient DNA samples are being included and compared to 70 modern samples.

National Geographic continues to publish research papers and have published 55 to date, with 5 more being near publication which is expected yet this year. I will publish a list shortly on this blog.

One of the questions that has been concerning genetic genealogists is how the recent sale of part of the National Geographic assets will affect the Genographhic project.
I asked Miguel privately, and he said that the research arm stays under the nonprofit National Geographic, but that the kit and website have both fallen in the group of products that have been sold to 21st Century Fox. Miguel said that he really didn’t have any answers at this time, but that the research continues and that the details are being worked out.
For those who don’t know, Spencer Wells stepped down as the director of the Genographic Project several months ago, but remains involved in a consulting capacity.
Michael Davila, Director of Product Marketing
Bennett introduced Michael Davila, the new Director of Product Marketing. Michael isn’t new to Family Tree DNA. He worked there for several years, from 2004-2011 when he left to work for a few years in the oil and gas industry, returning to FTDNA a few months ago in his new capacity.

Michael had one very short message. He knows there are problems and he is committed to getting them fixed and to providing tools for customers. The message, “Tools, tools, tools.”
Short, sweet and right to business.
I had a chance to meet with Michael outside of the conference room and I want to say that I’m very encouraged by Michael’s direct approach. He is insightful, understands the situation at hand and knows what needs to be done.
After Michael’s brief commentary, a general Q and A session followed. One of the questions was for Michael, and I just happened to catch this candid of Michael and Bennett! Not sure what Michael was saying, but it looks like it gave Bennett a migraine!

Actually, I think Bennett is concentrating on deciphering a question submitted by an attendee.
ISOGG Party
Saturday evening is traditionally the ISOGG party, but it’s not like a party you might generally think of. There is a lot of tutoring and collaborating that goes on. Friendships are made and renewed. Just being together is great. I mean, it’s not like we can have a discussion about SNP mutations rates at the dinner table at home.

You can see two different groups discussing aspects of genetic genealogy here.
At the party and also at Sunday’s sessions, lot of people were wearing the cool t-shirts gifted to participants by Family Tree DNA.


When we got back to our rooms, we discovered that even the hotel staff was in the spirit!!

ISOGG Meeting
Sunday morning is traditionally the ISOGG meeting. Not everyone attends, unfortunately, because not everyone is a member. Everyone is welcome, and since membership is free, it’s easy to join at www.isogg.org.

Katherine Borges was the original founder of ISOGG and still functions as the Director. ISOGG is celebrating its 10th anniversary this year, after being founded after the first genetic genealogy conference in Houston in 2004.

Katherine found a few photos of that first conference which was only one day and was held in a facility later destroyed by hurricane Ike, I believe. You can see more 2004 photos here and photos from other years here.

I don’t think Max (above) and Bennett had any idea what kind of a legacy they were creating with that first conference. History was being made.

Another function of ISOGG was the creation and maintenance of the Y SNP Tree. The tree was begun 9 years ago and has been organized and maintained by Alice Fairhurst this entire time.

In 2012, the Y tree had 800+ SNPs, but beginning with the introduction of the Big Y test, the SNP tsunami began. Today, there are over 15,000 SNPs on the tree, all entered by hand by Alice. Fortunately, each haplogroup has a coordinator, but still the increase in SNPs and the magnitude of the task at hand has been overwhelming.
Quality has to be maintained, because the tree is regularly referenced by academics as well as by genetic genealogists. Today, any SNP found in a Big Y type of next generation scan test must but be confirmed by Sanger sequencing. I know this is frustrating to some, but given the uncertainty of scanned SNPs, it’s also essential to maintaining the tree’s integrity.
Alice recently retired from heading the ISOGG Tree project and was presented with an award for her nine years of service to the genetic genealogy community by Katherine Borges on behalf of ISOGG.

In Alice’s comments, she said that “We have all driven a new industry.” Alice played a central and pivotal part.
Alice received a richly deserved standing ovation.

But we weren’t the only ones thanking Alice. Max and Bennett presented Alice with a certificate of appreciation for her years of service as well.

I was really pleased to see this. Not only is Alice extremely deserving of the recognition, but volunteers are too often unthanked and under-appreciated.
Panel Discussion: Protecting Ourselves: AHGS and Genetic Genealogy Standards
Have you ever been invited to a party, had to decline with genuine regrets, then later, been very glad that you didn’t attend. This describes the AHGS meetings for me. In 2013, I was invited to the first conference and couldn’t attend, but given what transpired and the difficult environment at the conferences, I’m grateful in retrospect.
Those who did attend, and those who subsequently developed the Genetic Genealogy Standards document formed a panel, moderated by Bennett Greenspan, to discuss those meetings and standards.

Panel members left to right, Katherine Borges, Steven Perkins, Dr. Tim Janzen, Jennifer Zinck and Debbie Parker Wayne.
In a nutshell, genetic genealogy had come to the attention of the American Human Genetics Society and not in a positive way. They didn’t understand what we are doing, and they became somewhat polarized on the idea of “harms.” What harms, you ask? Well, so did we. Apparently the harms they are concerned about are things like Y DNA testing revealing non-parental events.
The good news is that after this years meeting, it appears that the word “harms” has been removed and as a proactive measure, the genetic genealogy community created its own standards and guidelines.
You can read the resulting standards created by genetic genealogists here.
Brad Larkin Presents Genetic Genealogist of the Year Award
Brad Larkin, on behalf of the Surname DNA Journal, presented the second annual Genetic Genealogist of the Year Award to Maurice Gleeson.

The Surname DNA Journal would be a wonderful resource to publish many of these presentations – hint, hint to the presenters!!!

Maurice has organized Genetic Genealogy Ireland in Dublin since 2012, in addition to presenting widely. He also has a very successful series of genetic genealogy videos on YouTube which I highly recommend.

Maurice is a man of many talents – a psychiatrist and pharmaceutical physician, a professional actor and of course, a genetic genealogist. He has another talent as well – he can make absolutely anything interesting. If you ever hear that Maurice is giving a lecture on dust – by all means attend! It will be the highlight of your week, I guarantee.
Maurice, congratulations on your well-deserved honor and Brad, thank you for recognizing one of our colleagues.
Maurice Gleeson – Combining SNPs, STRs and Genealogy to Build a Surname Origins Tree

Maurice, like many of us, wants to be able to use STRs and SNPs in combination with genealogy records to construct accurate lineage trees. In addition, when genealogy records connecting people to their common ancestor are missing, we’d still like to be able to construct at least a hypothetical or genetically accurate tree.

During this process, Maurice encountered several challenges, including.
- Parallel mutations
- Back mutations
- Markers behaving unusually
- Multi-copy markers
- Unstable markers
- Lack of mutation rate for some markers
- Lack of standardized mutation rate for SNPs
- Difficulty determining if a SNP is present or absent
- Convergence issues
- False negatives
- False positives
- Unregulated naming
- Project members testing at different levels
Maurice did, however, provide us with the secret to success.

He began with hot cocoa and chocolate, but he said by the end of the project, he had an empty whiskey bottle and was taking anxiety medication:)
Maurice began by drawing a typical pedigree chart, based upon the project results, reflecting what he believed would be where the mutations would have occurred based on line marker mutations.

Then, with the assistance of Ralph Taylor, he drew Fluxus diagrams of the likely joining patterns of each set of possible outcomes. The outcomes included and then omitted various markers experimentally for various reasons related to the challenge list above.

Maurice shared lots of slides with us reflecting several different mutation sequence possibilities. I have omitted them, in part because I can’t explain why Maurice did what he did. I understood it at the time, but without the slides to take notes on, I don’t think I could reconvey it correctly. I would suggest that you obtain his slides from this link and view those in conjunction with Jennifer Zinck’s notes from his lecture.
In the end, Maurice did reach a “most probable” fit for both STRs and SNPs, although with some caveats, some of which were caused by participants who had only tested at 37 markers.

Maurice closed with lessons learned and future opportunities.




Maurice, this would make a fantastic YouTube video!! It was a wonderful lecture.
James Irvine – Surname Projects, Some Fresh Ideas

James has been working with the Scots-Irish Irvine project for the past ten years.

James Irvine and Maurice Gleeson are both trying to achieve many of the same goals, but are using somewhat different methods.
James creates his own spreadsheets for his project members which include not only STR markers, but lineage defining SNPs as well. Furthermore, James utilizes weighting for each STR marker based on its mutation rate, something the main project spreadsheet does not take into consideration in the step-wise mutations. However, James feels the TIP calculations, which do take mutation rate into consideration are really quite accurate, based on his reconstructed pedigrees.

Unfortunately, it’s at this point that my camera battery died completely, so I don’t have any further photos of his presentation, but would encourage you to download his slides for yourself at this link.
James discussed the varying SNPs reported by different entities and compares the results. The Big Y from Family Tree DNA, the results as analyzed by the appropriate project administrator(s), by a third party entity and then what he found himself. The various analysis and what they considered to be valid SNPs varied significantly.
Which one is accurate, and why and how does this in reality affect what we can surmise of the genealogy and constructing family trees? We just don’t know yet – but we are working with what we have.
One thing he mentioned is that the 495 STRs extracted by third parties from Family Tree DNA Big Y files are not necessarily reliable – which of course calls into question the reliability of any STR extracted from a next generation sequenced file. This also confirms why Sanger sequencing is required for SNPs to be added to the ISOGG tree.
Break Out Sessions
Sunday’s breakout sessions once again included sessions that would hopefully appeal to a wide range of audiences.
Katherine Borges – Nulls: The Value of Nothing (Y-DNA)
Matt Dexter – Surveying Ancestry Using Autosomal DNA Results
Jim Brewster – Getting Started with GAP
I attended Matt’s session, capturing only one photo with my phone. My apologies.

Matt discussed the foundation principles of autosomal testing and analysis, and how adoptees use this technology to find their families.
Roberta Estes – Y-DNA to Autosomal Case Study – Kicking It Up a Notch

(Thank you Jennifer Zinck for permission to use this photo.)
Lots of folks have sleepy Y DNA projects and wonder what else can be discovered utilizing these core projects. I did too, so I decided to try and see what happened if I expanded the Crumley Y project to include autosomal.
We began with 4 men who were Y DNA descendants of James Crumley born in 1712. These men descended from two of James’ sons, John and William.
We began our transition from Y to Y+Autosomal by upgrading all 4 men to the Family Finder test. We then set about recruiting additional members including those who are not male and do not carry the Crumley surname today.

The response was quite surprising and we quickly had 50 members, about 30 of whom descended from those same two sons. However, the descendants of the sons are today 7 generations distant, so 6th cousins, at the closest generation. The furthest distant from each other were 8th cousins once removed.
This begged several questions.

While the prediction models suggested that they wouldn’t match, they did.

In essence, we began to reconstruct the genome of James Crumley through his descendants by creating a spreadsheet showing how each Crumley descendant matched each other Crumley descendant. We utilized the same tools that we use for our own autosomal comparisons, some in a slightly different way.

This shows an example of three match groups of James’s descendants where the blue son, John’s descendants, are matching to the green son, William’s descendants on portions of chromosome 1.

Because I’m a visual person, I wanted to reconstruct James and Catherine’s genome on their chromosomes, so I utilized Kitty Cooper’s tools in ways they really weren’t quite designed for. Normally they are used to place your ancestral segment on your own chromosomes. Here, I used them to map James descendants matching DNA onto “James” chromosomes.

We actually accomplished several things and made multiple discoveries, many of which were entirely unexpected. I showed what we had discovered in the Y only project contrasted to the Y+autosomal project.

Last, I discussed how to transition a project from Y only to Y+autosomal.

My slides are available at this link, and I will be writing a series of articles from this research to be published in the upcoming weeks on my blog.
Dr. Connie Bormans, Laboratory Director, Tom Richard, Customer Support Manager and Mike Alexander, Director of Engineering
All three of these individuals have extremely critical positions at Family Tree DNA, all with very specific challenges.
Connie discussed sequencing technology and the differences between the different types of technology utilized for different tests.
Tom talked about several things he has done in less than a year at Family Tree DNA to improve customer service – and it has improved greatly. I spoke with him offline as well, and he has lots of plans going forward.
It’s wonderful to see such capable and motivated management team members.

Me with Tom Richard. I love meeting the staff and seeing them each year. It makes communicating with someone you know the rest of the year much easier.
Mike Alexander comes from NASA and his motto is the famous Gene Kranz statement, “Failure is not an option.” For those who don’t know, Kranz was the flight director credited with saving the Apollo 13 crew.
Little did Mike know, a few years ago, I sat in that seat in the Johnson Space Center and I have a t-shirt with that very saying.

Needless to say, I am greatly encouraged by Mike’s NASA experience and believe it will serve him, and Family Tree DNA, very well. Because, well, failure is not an option.
The Sale
I don’t know if Max and Bennett ever meant for this to happen, but it’s become a tradition that they announce a sale of some type during or at the end of the conference. They closed this conference with the announcement that the Holiday Sale was beginning and would continue until the end of the year. You can read about the sale and exchange coupons here.
Thank you Max and Bennett!
In Summary
This was an absolutely wonderful conference. I so enjoyed renewing old friendships and meeting new people. I’m very glad to see younger people and new admins interested and involved as well, because they are the next generation that will push what we’ve viewed as the frontier into the mainstream.
One day, we really will be constructing and reconstructing ancestors. We may be able to see their faces, know the color of their eyes and perhaps some of their traits. In another five years, we’ll be doing things we can’t even imagine today, and we’ll be pushing yet another line in the ever-expanding frontier of genetic genealogy.
Like I said when I closed my session, it takes a village. A village of participants to test, a village of administrators organizing and analyzing results, and pushing the proverbial research envelope. And it requires advanced tools and the supportive and incubational environment provided by Family Tree DNA. Without any of those things, we would fail. Thankfully, we won’t.
Maurice Gleeson perhaps said it best in his closing, “Max, Bennett, without you, there is no us.”
We truly are a partnership!

______________________________________________________________
Disclosure
I receive a small contribution when you click on some of the links to vendors in my articles. This does NOT increase the price you pay but helps me to keep the lights on and this informational blog free for everyone. Please click on the links in the articles or to the vendors below if you are purchasing products or DNA testing.
Thank you so much.
DNA Purchases and Free Transfers
Genealogy Services
Genealogy Research


























































































































































